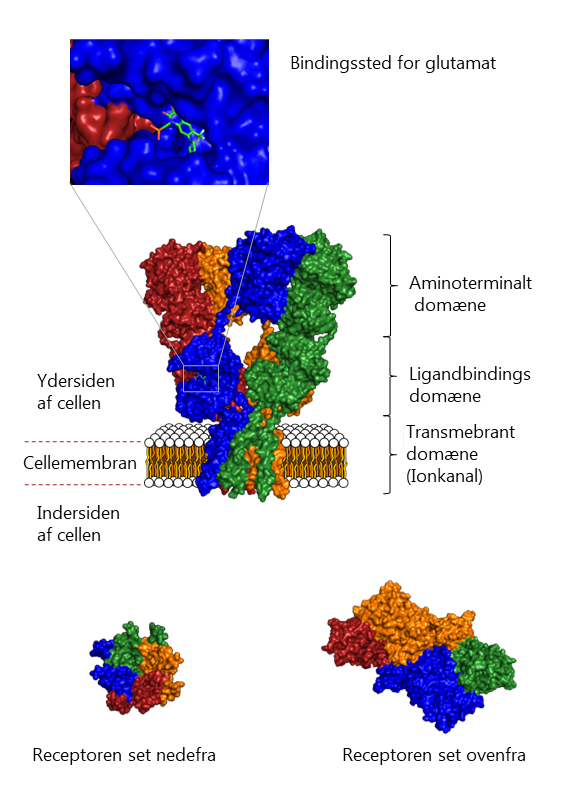
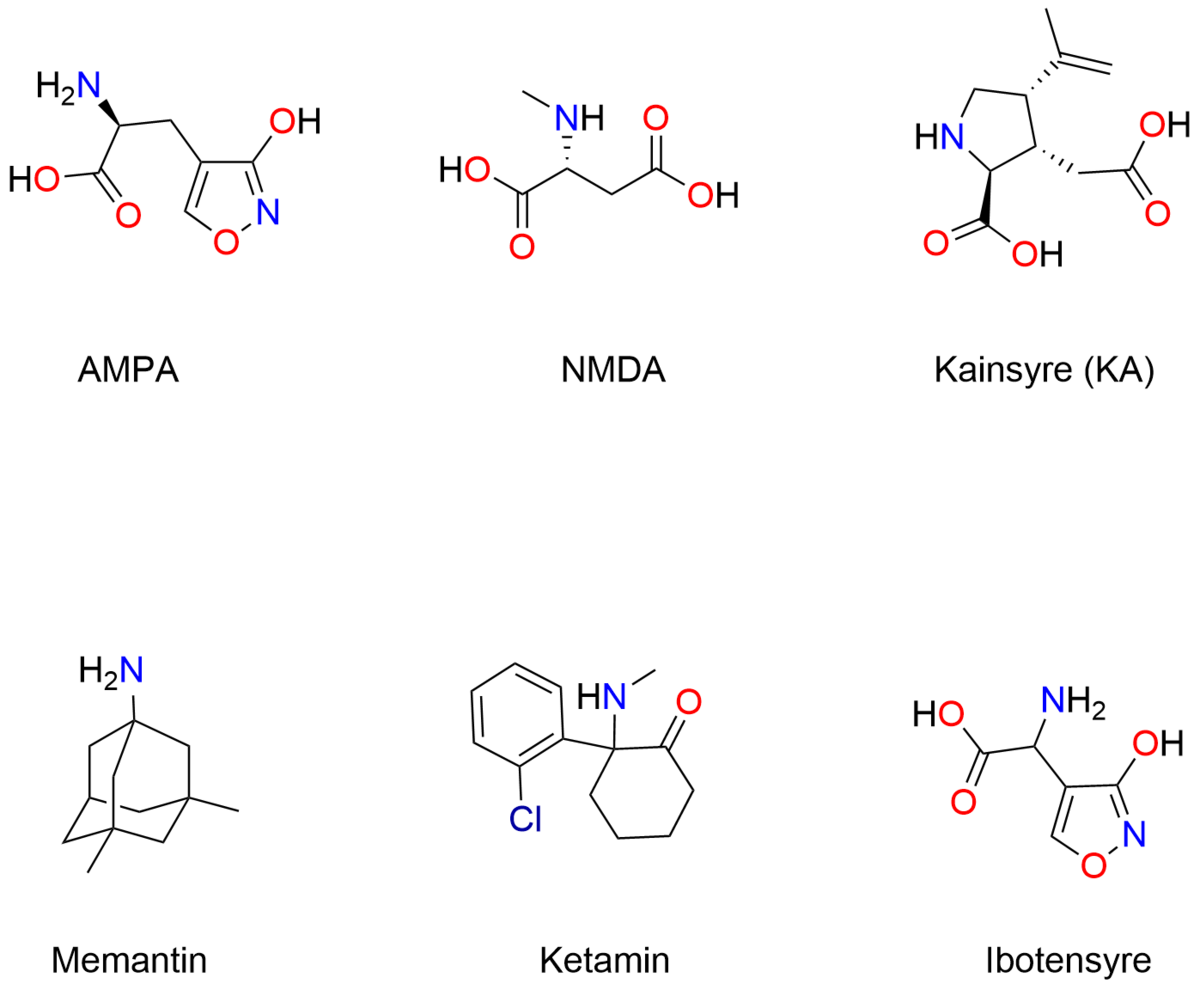
Udover den overordnede inddeling i to klasser kan glutamatreceptorerne yderligere underinddeles. De ionotrope glutamatreceptorer inddeles i 3 farmakologiske underklasser: AMPA-, NMDA- og Kainsyre-receptorer. Disse er faslagt efter, at receptorerne selektivt kan aktiveres af stofferne AMPA, NMDA og Kainsyre (Kainic Acid, KA). Derudover koder generne for endnu flere undertyper. Der er således klonet 4 AMPA-, 7 NMDA- og 5 KA-gener, der koder for de proteiner, der går sammen 4 af gangen for at danne receptoren. Da disse klonede undertyper også kombineres i de tetramere receptorer, er mangfoldigheden af glutamatreceptorer meget stor.
De mange undertyper har forskellige egenskaber, og spiller således forskellige roller i hjernen. For eksempler er NMDA-receptoren blokeret i ionkanalen af en magnesium-ion, som først forlader ionkanalen, når AMPA receptoren er aktiveret, og der er sket en delvis depolarisering af membranen. Endvidere kan kalcium-ioner passere gennem NMDA-receptoren, hvilket giver en større strøm. NMDA-receptorer menes at være involveret i hjernens hukommelse gennem en mekanisme, der kaldes langtidspotentiering (long term potentiation, forkortet LTP). Når NMDA-receptoren stimuleres gentagne gange, øges "styrken" af synapsen mellem de to nerveceller ved et den store strøm af kalcium-ioner fører til flere receptorer i og større "følsomhed" af synapsen.
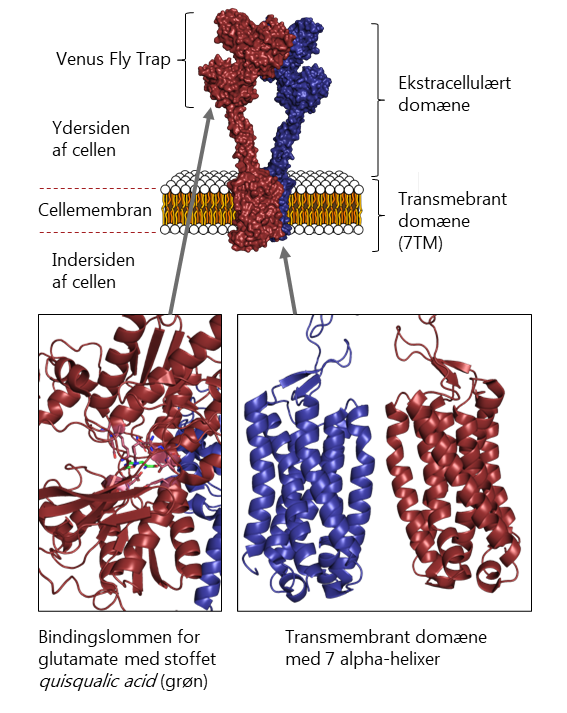
Også de metabotrope receptorer kommer i forskellige udgaver. I mennesker koder generne for 8 forskellige undertyper af metabotrope receptorer (mGluR1-8). Det signal, som metabotrope glutamatreceptorer overfører, kan virke både stimulerende og hæmmende afhængigt af hvilken undertype, der aktiveres. To af undertyperne, mGluR1 og mGluR5, er aktiverende og resten er hæmmende.


Kommentarer
Kommentarer til artiklen bliver synlige for alle. Undlad at skrive følsomme oplysninger, for eksempel sundhedsoplysninger. Fagansvarlig eller redaktør svarer, når de kan.
Du skal være logget ind for at kommentere.